Diploneis parma

-
Category
-
Length Range30 µm
-
Width Range20 µm
-
Striae in 10 µm14
-
ContributorSarah Spaulding - Jul 2019
-
ReviewerElena Jovanovska - Jun 2021
Identification
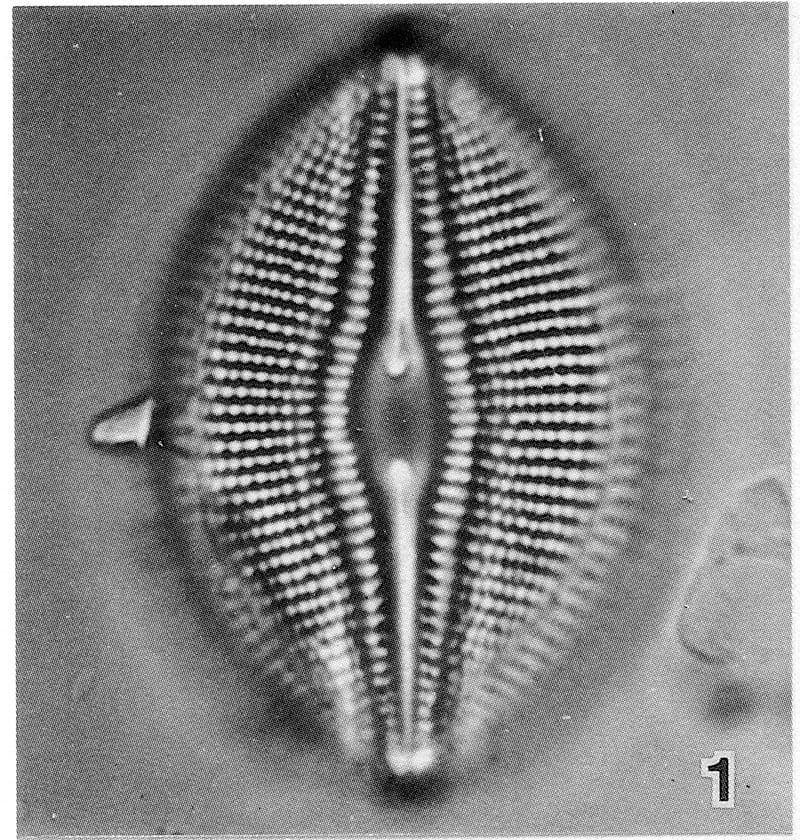
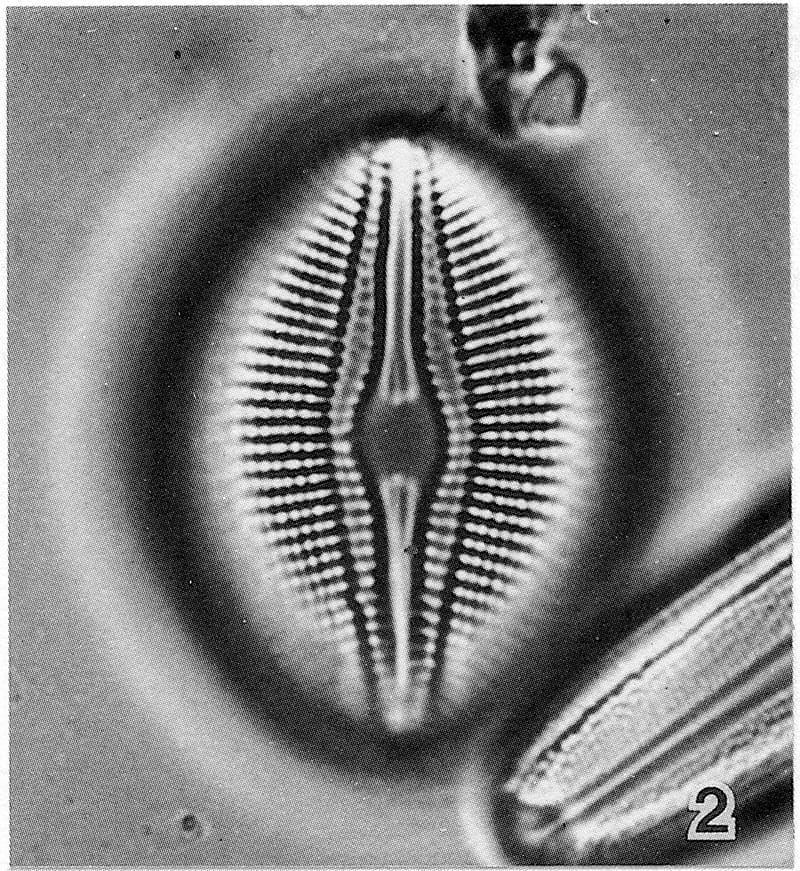
View image metadata
Type
Description
THIS TAXON HAS NOT BEEN VERIFIED IN NORTH AMERICA.
Diploneis parma, however, is frequently misidentified or confused with other species of Diploneis.
The purpose of including a treatment of a taxon in Diatoms of North America that is not verified is to alert practitioners to the taxonomic and nomenclatural issues related to D. parma.
Diploneis parma was originally described by Cleve (1891). Publication of the lectotype of D. parma (Idei and Kokayasi 1986) and subsequent work, shows a long history of misidentifications in the diatom literature. Misidentifications continue to the present day, as shown more recently (Lange-Bertalot and Fuhrmann 2017). According to these two publications, D. parma has only been verified from Finland, Russia and Japan.
The original description of D. parma includes three localities (Rosslängen, Smaland, Sweden; Lake Lojo, Ab. 36, Finland; Lake Viando, Sb. 5 Finland). The slide from Lake Lojo (No. 12:36, Regio aboënsis, Lojo) was designated as the lectotype. Only two specimens were found on this slide, both are reproduced (above) from Idei and Kobayashi (1986).
Since the original publication, Diploneis parma has been reported by a number of workers in Europe (Hustedt 1937, Mölder and Tynni 1973, Krammer and Lange-Bertalot 1986, Hofmann et al. 2011) and North America (Stoermer 1978). All of these reports, however, illustrate specimens that do not conform to the type of D. parma.
Observations of D. parma, as based on lectotype specimens (Lange-Bertalot and Fuhrmann 2017):
L: 26-46
W: 17-27
S: 12-13 in 10 (15-16 in Finland, 12-14 in Russia)
A: 12-13 in 10
"Valves broadly elliptical, sometimes weakly rhombic-elliptical. Ends obtusely, more or less cuneately rounded. Length 26-46 µm, breadth 17-27 µm; length-to-breadth ratio ca. 1.3-1.7, i.e. rather low and with low variance in comparison to other taxa of the genus. Raphe filiform, curved and together with the internal flanking depressions gradually expanding to the central ends. Distal ends appear indistinct in LM. Central area ca. 3-4.5 µm wide, variable in shape from narrowly elliptic to moderately broad-elliptic, sometimes slightly constricted dependent on focus. Appendices (horns) are present as proximal raphe ends protrude into the central area. Canal system lanceolate, ca. 6-8 µm broad proximally, marked by two, rarely three canal raphe areolae set off from the striae areolae. Striae radiate throughout, 15-16 in 10 µm in specimens from Finland and Germany while 12-14 in 10 µm in specimens from Lake Ladoga (Russia) and from Japan. Arolae may appear at first uniseriate in LM but can be resolved as opposedly biseriate with appropriate focusing, 18-20 in 10 µm. This can be confirmed by SEM and TEM observations (see Idei & Kobayasi 1988, pl. 1-3). The areolae become alternating biseriate in the marginal parts."
Autecology
According to Lange-Bertalot and Fuhrmann (2017), Diploneis parma is restricted to the northern boreal regions of Eurasia. In these few sites, it occurs infrequently and in low abundance.
Although it has frequently been recorded in North America, these records should be re-examined. The North American taxa likely belong to another species.
Please contact us with any records you suspect could belong Diploneis parma.
-
Size Range, µm3
-
Motility
-
Attachment
-
Habitat
-
Colony
-
Waterbody
- Learn more about this
Original Description
D. Parma Cl. n. sp. - Valve broadly elliptic, 0,03 m.m. in length and 0,02 m.m. in breadth. Central nodule quadrate. Furrows arcuate, equally bent, approximate to the prolongations of the central nodule. Costae 14 in 0,01 m.m.; puncta 20 in 0,01 m.m., forming longitudinal lines, crossing the costae.
This species has a great resemblance to the preceeding [Diploneis finnica] but is only half the size and has twice the number of costae and puncta in 0,01 m.m.
Pl.II, fig. 10.
I have found this form in the lake Rosslängen (Smaland, Sweden) and from the two finnish localities: Lojo Lake (Ab. 36) and Viando (Sb. 5), exactly similar in all these localities.
-
AuthorCleve 1891
-
Length Range30 µm
-
Width20 µm
-
Striae in 10µm14
Original Images


Citations & Links
Citations
-
Publication Link: 10.5507/fot.2015.027
-
Publication Link: 10.1016/S0380-1330(78)72181-7
Links
-
Index Nominum Algarum
Cite This Page
Spaulding, S. (2019). Diploneis parma. In Diatoms of North America. Retrieved April 03, 2026, from https://diatoms.org/species/61804/diploneis-parma
Responses
The 15 response plots show an environmental variable (x axis) against the relative abundance (y axis) of Diploneis parma from all the stream reaches where it was present. Note that the relative abundance scale is the same on each plot. Explanation of each environmental variable and units are as follows:
ELEVATION = stream reach elevation (meters)
STRAHLER = distribution plot of the Strahler Stream Order
SLOPE = stream reach gradient (degrees)
W1_HALL = an index that is a measure of streamside (riparian) human activity that ranges from 0 - 10, with a value of 0 indicating of minimal disturbance to a value of 10 indicating severe disturbance.
PHSTVL = pH measured in a sealed syringe sample (pH units)
log_COND = log concentration of specific conductivity (µS/cm)
log_PTL = log concentration of total phosphorus (µg/L)
log_NO3 = log concentration of nitrate (µeq/L)
log_DOC = log concentration of dissolved organic carbon (mg/L)
log_SIO2 = log concentration of silicon (mg/L)
log_NA = log concentration of sodium (µeq/L)
log_HCO3 = log concentration of the bicarbonate ion (µeq/L)
EMBED = percent of the stream substrate that is embedded by sand and fine sediment
log_TURBIDITY = log of turbidity, a measure of cloudiness of water, in nephelometric turbidity units (NTU).
DISTOT = an index of total human disturbance in the watershed that ranges from 1 - 100, with a value of 0 indicating of minimal disturbance to a value of 100 indicating severe disturbance.
Diploneis parma
- Length to breadth ratio 1.3-1.7
- Central area wide
- Proximal raphe ends protrude into central area
- Areolae biseriate
This taxon has not been verified in North America.
Valves are elliptic to rhombic-elliptic. The valves are relatively broad, with a length to breadth ratio of approximately 1.3-1.7. The central area is wide, 3-4.5 µm. According to Lange-Bertalot and Fuhrmann (2016) areolae typically appear to be uniseriate in LM, however, with critical...