





Credit: Rosalina Stancheva






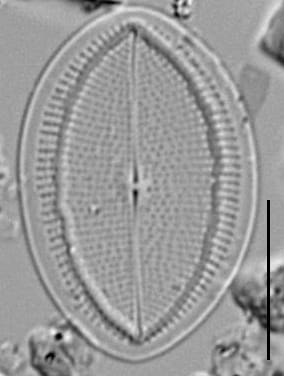
Valves are elliptic to round-elliptic with obtusely rounded apices. Valves are somewhat lopsided.
The raphe valve has a narrow axial area, a very small elliptic central area, and a straight to slightly deflected filiform raphe. Striae are slightly radiate at the center to strongly radiate and curved toward the apices. Areolae are 18-25 in 10 µm, clearly distinguishable with LM. Striae are interrupted by a hyaline submarginal ring with uneven margins separating one to ten submarginal areolae organized in longer rows in the middle, and shorter rows toward the apices. Valvocopula attached to raphe valve is open and irregularly finely fimbriated, but in smaller valves the fimbriae are weakly developed and often very short to see under LM.
The rapheless valve has striae that are slightly radiate at the center to strongly radiate and curved toward the apices. Fascia of 3–6 long striae, separated by short marginal striae not reaching the sternum. Areolae are short and dash-like, 13-20 in 10 µm, forming 8–21 undulating longitudinal lines per hemivalve and increasing in number with valve width. The sternum is narrow, linear and slightly deflected. Valvocopula attached to rapheless valve is open and smooth, but with fine irregularly denticulate advalvar rim, not always detectable with LM.
Cocconeis czarneckii belongs to the C. placentula species complex. Recent molecular studies of freshwater unialgal strains of Cocconeis (Jahn et al., 2009, 2020, Romero and Jahn 2013, Mora et al. 2021) set the baseline for defining species within this group typifying several well-known species that are extremely difficult to classify solely based on morphology.
The fine striation pattern of rapheless valve of C. czarneckii
is very similar to the illustrations of C. placentula (Patrick and Reimer 1966, Plate 15, Fig. 7) and C. placentula s.l. (finely punctate forms) (Lange-Bertalot et al. 2017, Plate 20, Figs 1, 2). However, the areolae on rapheless valves of C. placentula
and C. euglypta (as epitypified by Jahn et al. 2009 and Romero and Jahn 2013) are longer and form less longitudinal lines per hemivalve (maximum 6) in contrast to C. czarneckii. Cocconeis lineata and C. coreana have rapheless valves with high number longitudinal lines per hemivalve (9-12 in former, and 6-17 in later) similarly to C. czarneckii, but differ by their more elongate, elliptical valves, which are proportionally longer with more rounded apices. The areolae in C. coreana rapheless valves are larger and longer than in C. czarneckii, thus measuring only 9 to 12 in 10 µm. Raphe valves of C. lineata and C. coreana differ by less radiate striae (nearly parallel in the middle) and narrow submarginal area with one to three areolae in a row. Furthermore, the raphe valve valvocopula in C. lineata
is smooth, in C. coreana is fimbriate but with coarser clamp-like fimbriae. The largest valves of C. czarneckii are similar in areolae pattern to C. klamathensis and C. grovei. However, they have several differences from C. czarneckii, the most striking being the strongly arched valves of C. klamathensis and its raphe valve with branched fimbriae only in the middle; and the larger valve size and lower stria density in C. grovei.
This species was described and genes were sequenced from unialgal strain collected from the shoreline at the Iowa Lakeside Laboratory, West Lake Okoboji, Dickinson County, Iowa. Its geographical distribution and ecological preferences are unknown so far. This species is not characteristic for streams in California, although single valves have occasionally has been recorded.


Cocconeis czarneckii Stancheva, L.D.Mora and R. Jahn sp. nov.
Figs 1–76
DESCRIPTION BY LM (Figs 1–26): Valves elliptical to round elliptical, somewhat lopsided with obtusely rounded apices (Figs 1–17) to rounded in the largest cells (Figs 18–26). SV striae slightly radiating at the centre to strongly radiating and curved towards the apices; in fascia of 3–6 long striae, separated by short marginal striae not reaching the sternum (Figs 1, 3–9, 18–20). Areolae in a short dash-line, forming 8–21 undulating longitudinal lines per hemivalve increasing in number with valve width. Sternum narrow linear and slightly deflected (Figs 1, 4, 8, 18–20). Large and initial valves with a slightly diagonal sternum, separated by several areolae from the apices (Figs 18–20). RV with straight to slightly deflected filiform raphe, narrow axial area and very small elliptical central area (Figs 10–12, 14–17, 23–26). RV striae slightly radiating at the centre to strongly radiating and curved towards the apices, composed of small nearly round areolae, clearly distinguishable with LM in most specimens (Figs 11, 12, 16, 26), but more delicate and sometimes disordered near the centre (Figs 10, 14–17, 23–26). Striae interrupted near valve margin by thickened hyaline rim with uneven margins and isolated areolae visible in the largest valves (Fig. 26), separating a number of marginal areolae with longer rows in the middle of larger valves Figs 10–12, 14–17, 23–26). SV valvocopula open and smooth (Figs 2, 21). RV valvocopula open and irregularly finely fimbriated (Figs 13, 22, Figs 26); fimbriae in smaller specimens short.
DIMENSIONS: Length 17.7–51 μm; width 11.3–37.9 μm; SV striae: 20–26 in 10 μm with 13–20 areolae in 10 μm; SV striae forming 8–21 longitudinal rows of areolae per hemivalve; RV striae: 18–30 in 10 μm with 18–25 areolae in 10 μm. Because the strain auxosporulated during our investigation, the size range contains only small and large cells but no intermediate sizes. This means that immediate post initial cells were included in the above size ranges.
DESCRIPTION BY SEM (Figs 27–63): SV face flat to slightly convex with a short mantle (Figs 27–31, 35–37). Striae uniseriate, composed of dash- like areolae (Figs 27–31); interspersed short striae near mantle of one to six shorter areolae (Figs 29–31). External areolae caved in with narrow slit-like opening, occluded by lattice-structured hymen with slits (Figs 32–34). Internal areolae transversally elongated (Figs 35, 37) or roundish (Fig. 36). Sternum pronounced, linear, separated from valve apices by at least two – one long and one short – areolae in the smaller valves (Figs 28, 29, 37), and up to 10 in larger valves (Figs 30, 31). SV VC open, abvalvar side smooth and slightly angular concave (Fig. 40), advalvar side with an irregular denticulate rim (Figs 38, 39, 45), which connects with valve (Figs 43, 44). First copula open (Figs 41–43) with smooth convex abvalvar side (Fig. 42), and a regular undulate rim on the advalvar side (Fig. 41), which fits the advalvar surface of SV VC (Fig. 40). RV face flat with a weakly concave mantle (Figs 47–49, 53– 57). Striae uniseriate (Figs 47–49, 53–57), composed of irregular variably sized mostly round areolae (Figs 51, 62) to elongate in larger valves (Fig. 63), regularly spaced (Figs 48, 49, 61) or disorderly (Figs 51, 52, 58). The first areolae of each stria are very close and orderly at the raphe whereas the other areolae of the striae are irregular (Figs 47, 48, 53, 56). Areolae internally closed by lattice-like hymenes (Figs 50–52), visible also externally (Figs 50, 59). Internally, striae interrupted by a raised submarginal hyaline rim separating one to three submarginal areolae near the apex (Figs 49, 53–55, 57), and up to five areolae in the middle in smaller specimens (Figs 49, 52), up to 10 in larger specimens (Figs 47, 48); the numbers depend on the size of the valve with the larger valves having more areolae at the centre as well as at the apices. Submarginal areolae either showing a gap at the apices (Figs 49, 53), a single areola (Fig. 54) or several areolae may circumradiate the apices as seen in some small (Figs 55, 57), and larger specimens (Figs 47, 48, 56); nevertheless, the number of submarginal areolae is always lower at the apices than in the middle. Externally, the hyaline rim in larger valves interrupted by some areolae of the valve and submarginal striae (Fig. 56); in smaller valves the hyaline rim may be invisible (Figs 53, 54), partially visible with dispersed areolae (Fig. 55) or well defined (Fig. 57). Raphe filiform, internal proximal raphe fissures deflected to opposite sides and distal raphe endings with visible helictoglossae at the hyaline marginal rim (Figs 47–49). External proximal raphe ends drop-shaped, distal ends straight and distant from margin (Figs 53–57). RV valvocopula open, fimbriated (Figs 60–63); the fimbriae short and irregular becoming unnoticeable in some valvocopulae (Fig. 60), but well-developed, laterally expanded in other (Figs 61–63). The fimbriae are void of any other morphological structure such as papillae.
LIVING CELLS AND AUXOSPORES (Figs 64–76): Twenty days after receiving the strain from UTEX in Austin, Texas, and continuing culturing at CSUSM in San Marcos, Southern California, we observed and documented auxospore formation (Fig. 68) and immediate post initial cells (Figs 64–66, 69), as well as other living cells all with protoplasts (Figs 67, 70–76). It is interesting to notice that the gap in the open copulae was distinguishable in the living cells with appropriate focus (Figs 64, 65, 73, 76). Twenty days after receiving the strain from UTEX in Austin, Texas, and continuing culturing at CSUSM in San Marcos, Southern California, we observed and documented auxospore formation (Fig. 68) and immediate post initial cells (Figs 64–66, 69), as well as other living cells all with protoplasts (Figs 67, 70–76). It is interesting to notice that the gap in the open copulae was distinguishable in the living cells with appropriate focus (Figs 64, 65, 73, 76).
HOLOTYPE: Slide ANSP GC65344, deposited at the Diatom Herbarium, Academy of Natural Sciences, Philadelphia, USA. Figure 7 represents the holotype. Based on the living strain UTEX FD23.
ISOTYPE: Slide B 40 0045143, Botanic Garden and Botanical Museum Berlin, Germany.

Stancheva, R. (2021). Cocconeis czarneckii. In Diatoms of North America. Retrieved July 22, 2026, from https://diatoms.org/species/206909/cocconeis-czarneckii
The 15 response plots show an environmental variable (x axis) against the relative abundance (y axis) of Cocconeis czarneckii from all the stream reaches where it was present. Note that the relative abundance scale is the same on each plot. Explanation of each environmental variable and units are as follows:
ELEVATION = stream reach elevation (meters)
STRAHLER = distribution plot of the Strahler Stream Order
SLOPE = stream reach gradient (degrees)
W1_HALL = an index that is a measure of streamside (riparian) human activity that ranges from 0 - 10, with a value of 0 indicating of minimal disturbance to a value of 10 indicating severe disturbance.
PHSTVL = pH measured in a sealed syringe sample (pH units)
log_COND = log concentration of specific conductivity (µS/cm)
log_PTL = log concentration of total phosphorus (µg/L)
log_NO3 = log concentration of nitrate (µeq/L)
log_DOC = log concentration of dissolved organic carbon (mg/L)
log_SIO2 = log concentration of silicon (mg/L)
log_NA = log concentration of sodium (µeq/L)
log_HCO3 = log concentration of the bicarbonate ion (µeq/L)
EMBED = percent of the stream substrate that is embedded by sand and fine sediment
log_TURBIDITY = log of turbidity, a measure of cloudiness of water, in nephelometric turbidity units (NTU).
DISTOT = an index of total human disturbance in the watershed that ranges from 1 - 100, with a value of 0 indicating of minimal disturbance to a value of 100 indicating severe disturbance.