Cocconeis coreana

-
Category
-
Length Range21-58 µm
-
Width Range11.5-36.6 µm
-
Striae in 10 µm18-24 raphe valve, 18-24 rapheless valve
-
ContributorRosalina Stancheva - Jan 2022
-
ReviewerSarah Spaulding - Feb 2023
Identification









Description
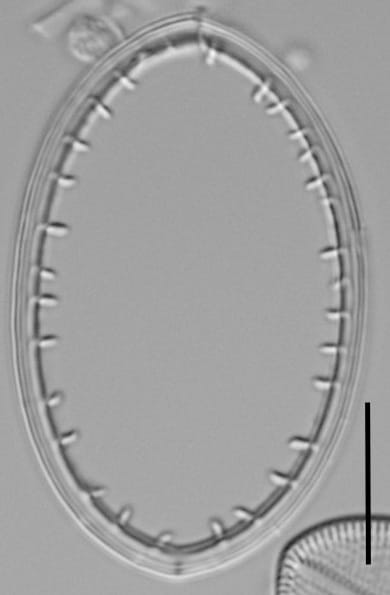
Valves are slender elliptic to elongate-elliptic. In some rare specimens, only rarely round-elliptic with obtusely rounded apices that are somewhat lopsided. The raphe valve has a narrow axial area, pronounced central area, and straight filiform raphe. Striae are parallel at the center and radiate and slightly curved near the apices. Areolae are 15-24 in 10 µm, and clearly distinguishable with LM. Striae are interrupted by a hyaline, submarginal ring with uneven margins separating 2 to 4 submarginal areolae. Valvocopula is open, with distinct, irregularly spaced, clamp-like fimbriae, clearly visible under LM.
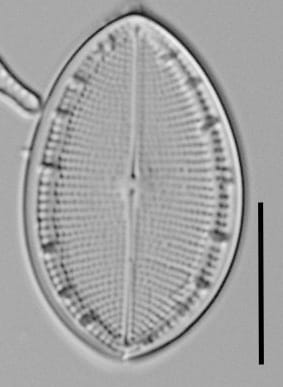
Striae of the rapheless valve are parallel at the center and radiate and slightly curved near the apices. Fascia are between 2-5 long striae, separated by short marginal striae not reaching the sternum. Areolae are prominent, dash-like, 11-14 in 10 µm, and form 6-17 longitudinal lines. The sternum is narrow, linear, sometimes widened near the center. Valvocopula attached to rapheless valve is open and smooth, but with irregularly denticulate advalvar rim, typically detectable with LM.
Cocconeis coreana is described and illustrated here from a single population, where auxospores were observed. This species from streams in California has rapheless valves with variable areolae length, which are slightly more dense than the original description.
Cocconeis coreana belongs to Cocconeis placentula species complex. Recent molecular studies of freshwater unialgal strains of Cocconeis (Jahn et al., 2009, 2020, Romero and Jahn 2013, Mora et al. 2021) set the baseline for defining species within this group typifying several well-known species that are difficult to classify solely based on morphology.
Morphologically similar species to C. coreana had been identified as C. placentula var. lineata by Patrick and Reimer 1966 (Plate 15, Figs 5, 6). Cocconeis coreana is morphologically most similar to C. lineata (as epitypified by Romero and Jahn 2013), which differ by its smaller finer frustules, rapheless valve with delicate and denser areolae (12-26 in 10 µm), raphe valve with more parallel striae and smooth valvocopula. Cocconeis placentula and C. euglypta (as epitypified by Jahn et al. 2009 and Romero and Jahn 2013) differ by rapheless valves with areolae organized in less longitudinal lines per hemivalve (maximum 6).
Cocconeis czarneckii differs by its proportionally wider valves, more radiate striae on both valves, rapheless valve with finer and denser areolae (13-20 in 10 µm), raphe valve with more submarginal areolae, forming longer rows in the middle of the valves, and valvocopula with more delicate fimbriae.
Autecology
Cocconeis coreana was described and genes were sequenced from unialgal strains collected from the Hyangsan waterfalls, Myohyangsan, DPR Korea. It seems this species is characteristic for running waters, because it is common and locally abundant in streams in California, particularly in the central and northern parts of the state. However, C. coreana is not confirmed genetically in California, so far.
-
Size Range, µm3
-
Motility
-
Attachment
-
Habitat
-
Colony
-
Occurrence
-
Waterbody
- Learn more about this
Original Description
Cocconeis coreana R. Jahn & B. M. Suh sp. nov.
Holotype. Slide B 40 0041501a, Botanic Garden and Botanical Museum Berlin, from the strain D119_029. Fig. 2F (SV; Fig. 2B RV) represents the holotype. SEM-stub deposited as B 40 0041501b. For molecular material and data see “Materials and Methods” and Supplementary Table S1.
Type locality. DPR Korea, Myohyangsan, Hyangsanwaterfalls, Sep 17, 2014, collected by Byung Moon Suh & Regine Jahn.
Registration. http://phycobank.org/102187.
Etymology. We are referring to the entire Korean Peninsula since this taxon was found in clean freshwaters in North and South Korea.
Description by LM (type strain only, Fig. 2). Valve outline is generally slender elliptical (Fig. 2A) but sometimes round elliptical (Fig. 2B), apices are obtusely rounded (length [AA]: 25.5-59.3 µm; width [TA]: 14.1-36.6 µm [n = 34]). The number of striae on both valves are similar but the individual punctae are different. The RV shows punctate striae with 18-24 rows per 10 µm and 15-24 areolae in 10 µm. The SV shows short dash-like striae with 18-24 rows per 10 µm and 9-12 areolae per 10 µm. The striae on both valves are parallel at the center and radiate near the apices; the striae of the SV seem to be in bundles of 2-5 striae (Fig. 2E-H). The RV shows a pronounced round central area (Fig. 2A-D). Only the RV has a rim of several rows of areolae at the valve mantle as a continuation of the striae separated by a submarginal hyaline rim (for details see SEM). In addition, the RV has a conspicuous valvocopula with irregular spaced clamp-like fimbriae (for details see SEM). Raphe is filiform and straight, axial area very narrow; sternum axial area very narrow. Concerning alive cells, in one strain we documented a single c-shaped plastid with serrated edges and a pyrenoid (Fig. 1A-C) and in two strains we observed auxospore formation (Fig. 1D-G) (not for the type strain).
Description by SEM (Figs 7-11). RV: The valve face is flat with a concave mantle separated by a pronounced (Fig. 8A & C). Striae are made up of round to slightly elliptical uniseriate areolae which are small (about 0.25 µm) and regularly spaced, increasing in size from the axial area towards the valve margin and the apices (Fig. 7E), forming a pronounced central area of about 6 rows of areolae length (AA) and 3 areolae wide on each side of the raphe ends (TA) (Fig. 7A-C). Internally, the areolae are occluded by round lattice-structured hymenes with short slits around the margin and tiny perforations in the middle (Figs 7D, 8B & D). On the mantle, the uniseriate valve face striae continue as uniseriate rows of two to four, generally three areolae (Fig. 7A-C); at each pole there is a gap of these mantle areolae (Fig. 7E). Externally, the raphe is straight, filiform, proximal raphe endings are slightly drop-like expanded (Fig. 7A-C). Internally, the proximal raphe fissures are bent to opposite sides (Fig. 8A & C); distal raphe endings are straight and end in weakly elevated helictoglossae on the hyaline rim (Fig. 8A & C). Valvocopula is open; long, slender and heavily silicified clamp-like fimbriae (without papillae and furrows) seem to be attached to the hyaline rim and are irregularly spaced at every 4th to 7th stria, or 3 to 5 clamps per 10 µm (Figs 8A & 10F).
SV: The valves are thicker than the RV (compare Figs 7D & 9G). The valve face is flat to slightly convex with a short mantle (Fig. 9A). Striae are made up of dash-like regular uniseriate areolae, undifferentiated unto the valve edge (Fig. 10A & D); internally they are rounded (Fig. 9D & F) whereas externally they are slit-like of different openness (Fig. 10B & E). Areolae are occluded by elongated lattice structured hymenes of about 0.7 µm length by 0.13 µm width with marginal slits and tiny holes in the center; the hymenes lie towards the outer side but not completely externally (Figs 9E, G & 10B). The sternum is very thin and sometimes shows the mark of a ghost raphe (Fig. 10A). The valvocopula is open with no visible fimbriae (Figs 9B & 10C; the fimbriae and valvocopula seen in Fig. 9C are from the RV). We also observed auxospore formation in SEM (Fig. 11) with remnants of incunabula (Fig. 11A & B) but apparently no perizonium.
Comparison to related taxa. With its largest length of almost 60 µm and width of up to a 43 µm C. coreana is generally bigger than C. placentula var. placentula (sec. Jahn et al. 2009, figs 28-43). Nevertheless, their length to width ratios overlap, with C. coreana having a broader ratio (see morphometric values in Table 1). The RV of C. coreana looks like a replica of C. placentula with generally similar numbers of striae and areolae, becoming larger towards the pronounced hyaline rim; also the numbers of areolae of the mantle below the hyaline rim are similar with about 3 (2-4 for C. coreana and 3-4 for C. placentula) per row but there is a gap at both poles in C. coreana (Fig. 7A-C) in contrast to continuing areolae in C. placentula (Jahn et al. 2009, figs 37 & 39). Further, in C. coreana the numbers and spacing of the longer clamp-like fimbriae of the raphe valvocopula are similar but there seem to be no small fimbriae in C. coreana (Figs 8A & 9C) in contrast to C. placentula (Jahn et al. 2009, fig. 42). The size of the central area around the drop-like proximal raphe ends is pronouncedly bigger in C. coreana (Fig. 7A-C) than in C. placentula (Jahn et al. 2009, figs 37 & 39). As expected for the genus Cocconeis, the biggest differences are found in the SV concerning numbers of the dash-like striae, size of the areolae and size of the sternum: C. coreana has a narrow sternum and closely spaced areolae of the size 0.6 µm by 0.2 µm externally (Fig. 10C); C. placentula has a broad sternum and widely spaced long and very thin areolae of the size of up to 1.5 µm by 0.05 µm externally (Jahn et al. 2009, figs 40 & 41).
Comparing C. coreana to C. lineata (sec. Romero and Jahn 2013, figs 2-8) is problematic since the only available data is from one strain without auxospores. C. lineata has a higher length to width ratio (see morphometric values in Table 1) which means that C. lineata is smaller and slenderer in comparison to C. coreana. The mantle of the RV of C. lineata shows 2-3 rows of areolae with a gap at the poles as C. coreana. The length to width ratios and the numbers and density of the SV areolae are similar in C. coreana and C. euglypta (sec. Romero and Jahn 2013, figs 9-18) (see morphometric values in Table 1). In C. euglypta the hyaline rim is barely visible externally and the raphe mantle areolae rows are only 2 in C. euglypta.
With 2.6-2.87% for rbcL and 6.1% for 18SV4 C. coreana has the lowest p-distance with the strain UTEX FD23 named C. placentula, but 4.0-4.1% for rbcL and 7.4% for 18SV4 to the epitype of C. placentula var. placentula (see detailed values in Tables 2 & 3).
-
AuthorR.Jahn and B.M.Suh 2020
-
Length Range25.5-59.3 µm
-
Width14.1-36.6 µm
-
Striae in 10µm18-24 raphe valve, 18-24 rapheless valve
Original Images


Citations & Links
Citations
-
Publication Link: 10.4490/algae.2020.35.8.1
-
Publication Link: 10.1080/00318884.2021.2003684
-
Publication Link: 10.1080/0269249X.2013.770801
Links
-
DiatomBase
Cite This Page
Stancheva, R. (2022). Cocconeis coreana. In Diatoms of North America. Retrieved July 22, 2026, from https://diatoms.org/species/206992/cocconeis-coreana
Responses
The 15 response plots show an environmental variable (x axis) against the relative abundance (y axis) of Cocconeis coreana from all the stream reaches where it was present. Note that the relative abundance scale is the same on each plot. Explanation of each environmental variable and units are as follows:
ELEVATION = stream reach elevation (meters)
STRAHLER = distribution plot of the Strahler Stream Order
SLOPE = stream reach gradient (degrees)
W1_HALL = an index that is a measure of streamside (riparian) human activity that ranges from 0 - 10, with a value of 0 indicating of minimal disturbance to a value of 10 indicating severe disturbance.
PHSTVL = pH measured in a sealed syringe sample (pH units)
log_COND = log concentration of specific conductivity (µS/cm)
log_PTL = log concentration of total phosphorus (µg/L)
log_NO3 = log concentration of nitrate (µeq/L)
log_DOC = log concentration of dissolved organic carbon (mg/L)
log_SIO2 = log concentration of silicon (mg/L)
log_NA = log concentration of sodium (µeq/L)
log_HCO3 = log concentration of the bicarbonate ion (µeq/L)
EMBED = percent of the stream substrate that is embedded by sand and fine sediment
log_TURBIDITY = log of turbidity, a measure of cloudiness of water, in nephelometric turbidity units (NTU).
DISTOT = an index of total human disturbance in the watershed that ranges from 1 - 100, with a value of 0 indicating of minimal disturbance to a value of 100 indicating severe disturbance.
Cocconeis coreana
- Raphe valve with striae interrupted by submarginal, hyaline ring
- Raphe valve with 15-24 areolae in 10 µm
- Valvocopula with clamp-like fimbriae
- Rapheless valve with striae parallel at the center
- Rapheless valve with 11-14 areolae in 10 µm
The raphe valve has a narrow axial area, pronounced central area, and straight filiform raphe. Striae are parallel at the center and radiate and slightly curved near the apices. Areolae are 15-24 in 10 µm, within a stria. Striae are interrupted by a hyaline, submarginal ring. Valvocopula with clamp-like fimbriae.
The rapheless valve has...