
Credit: L. Stratton
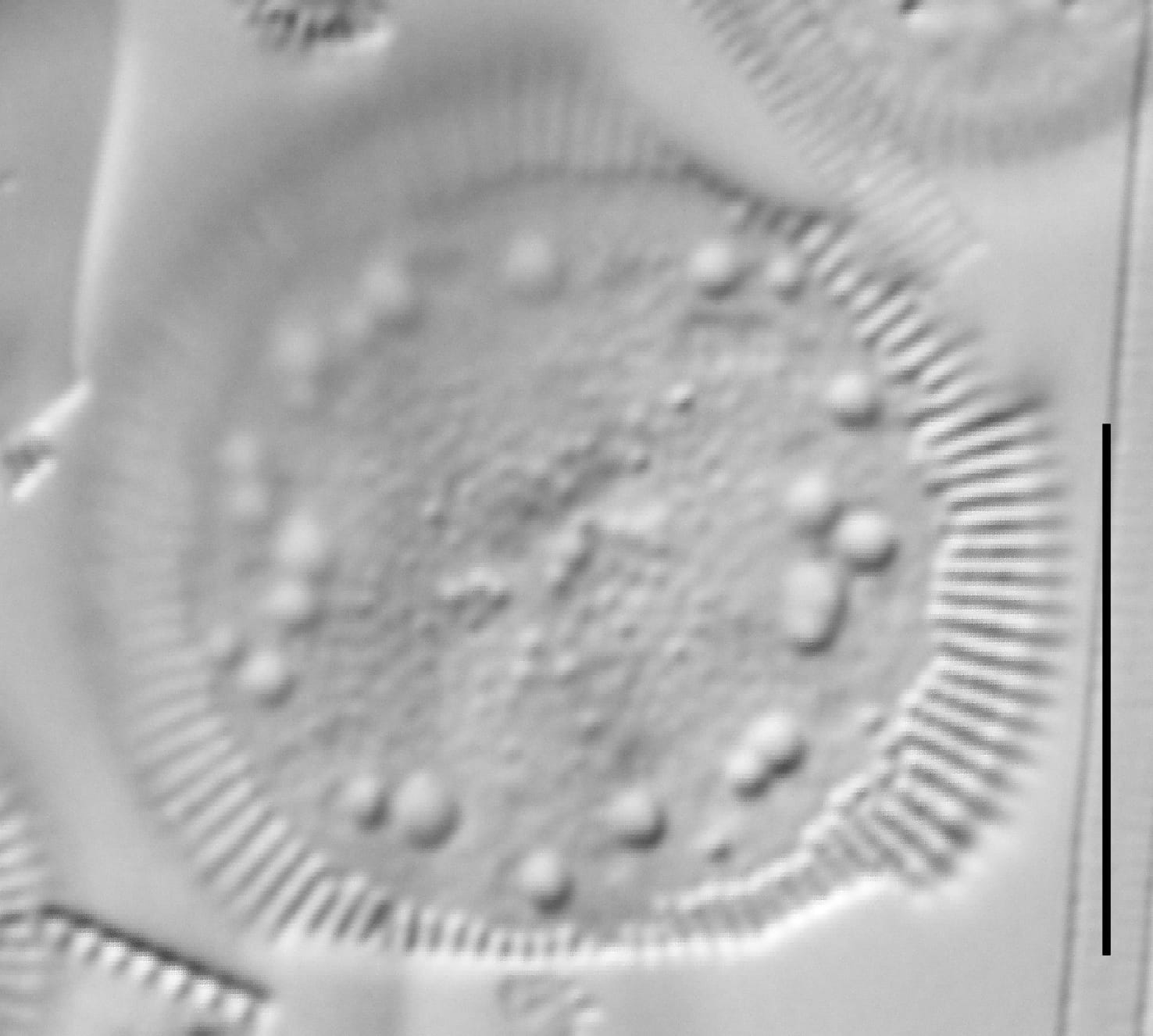
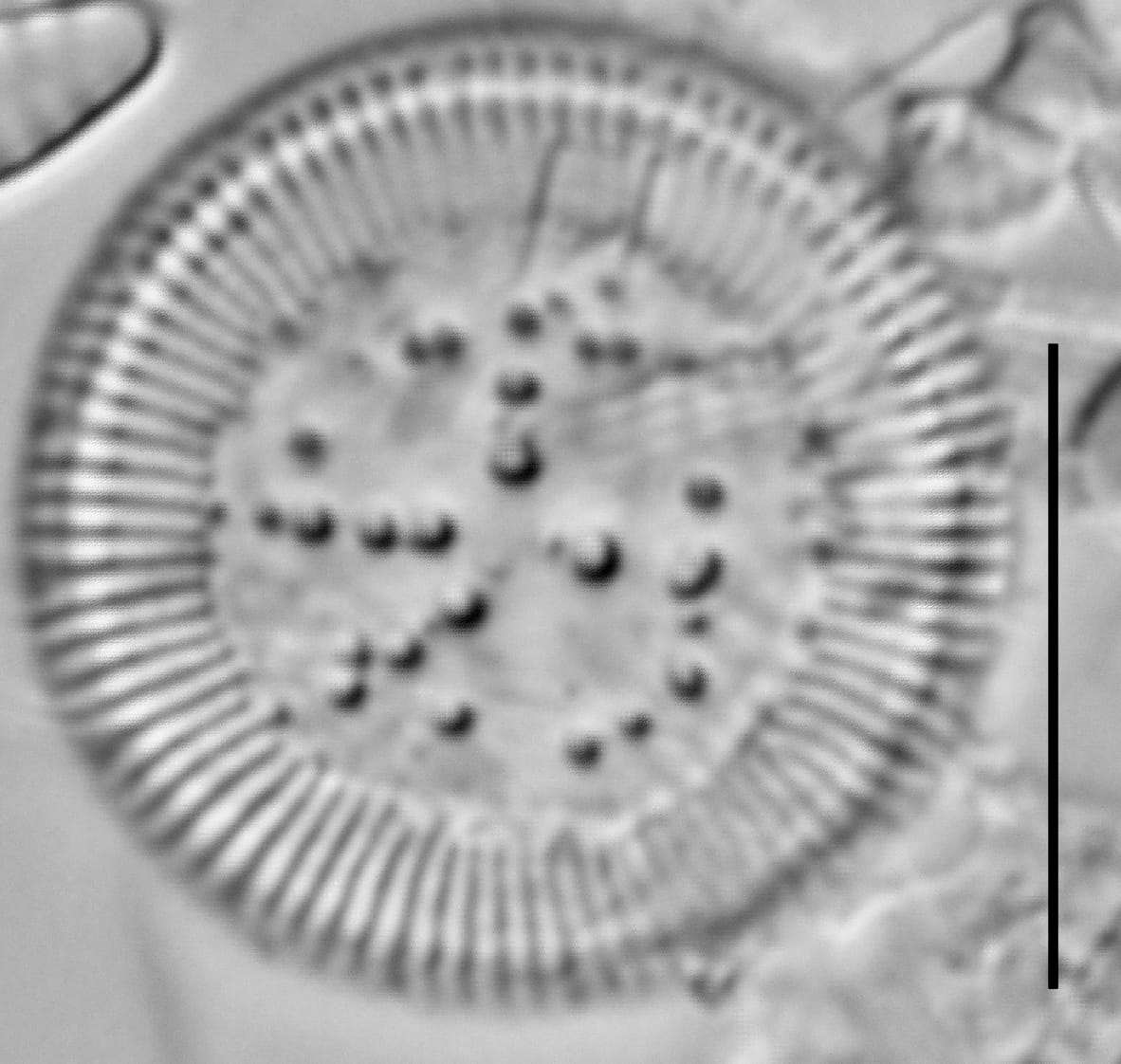
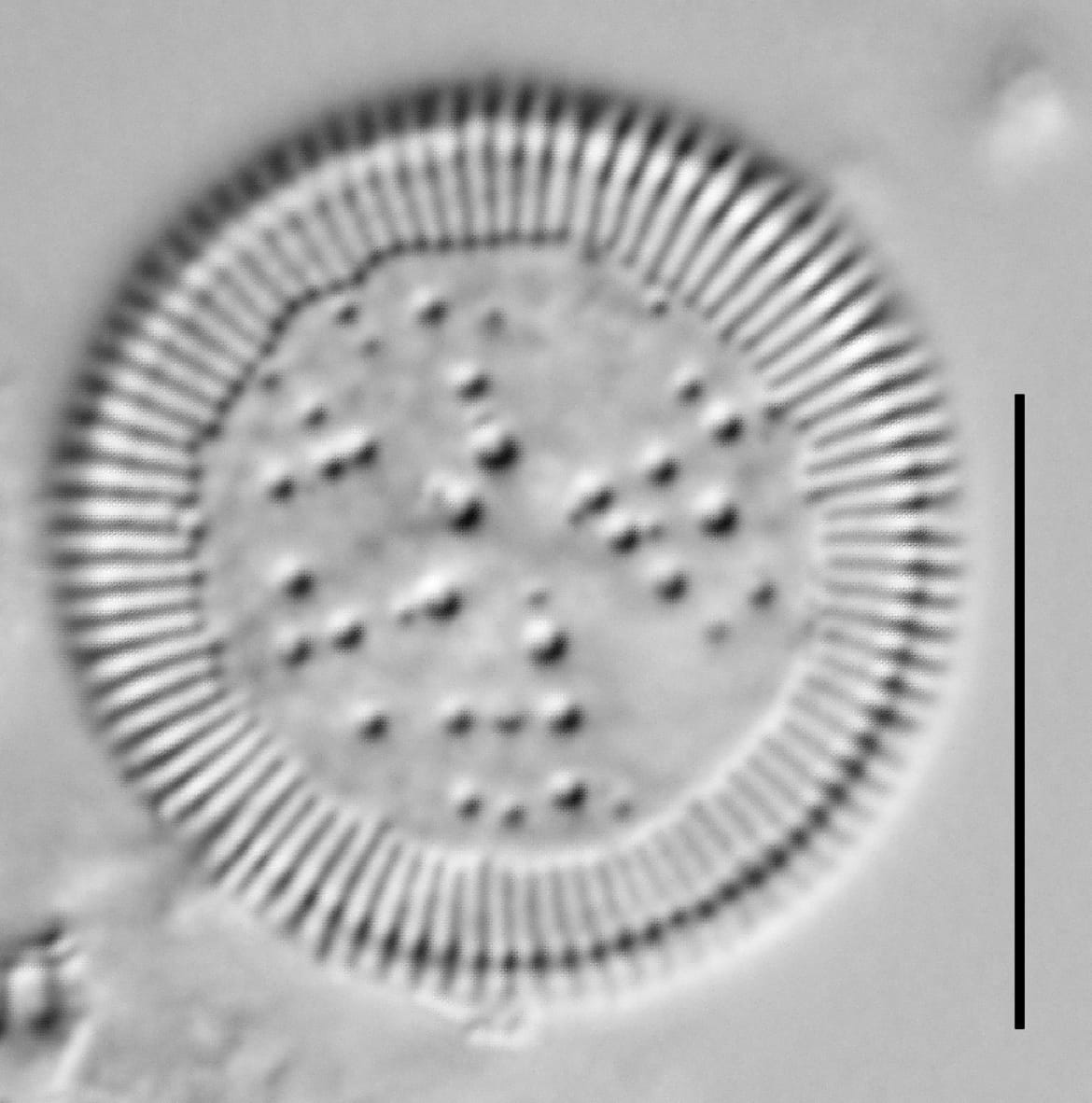
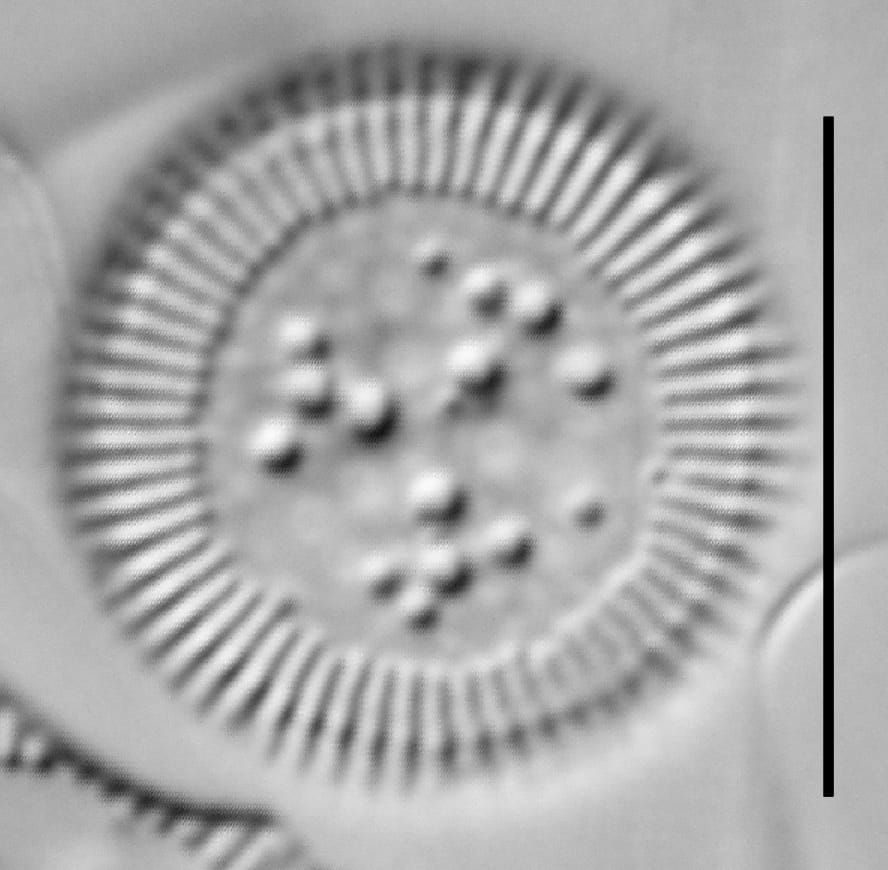
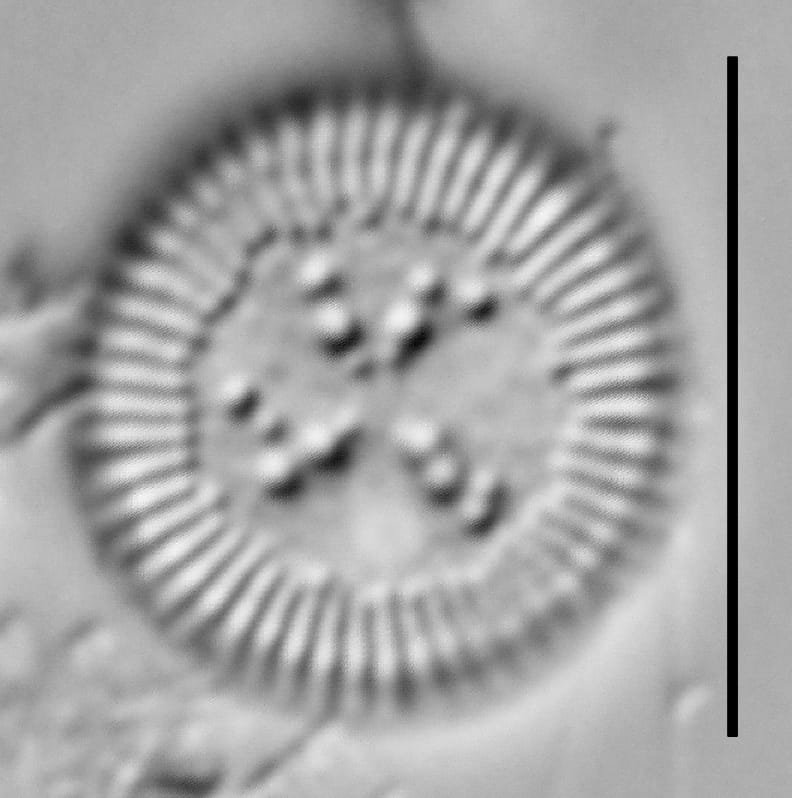
Cylotella rossii initial valve, Fallen Leaf Lake, CA.

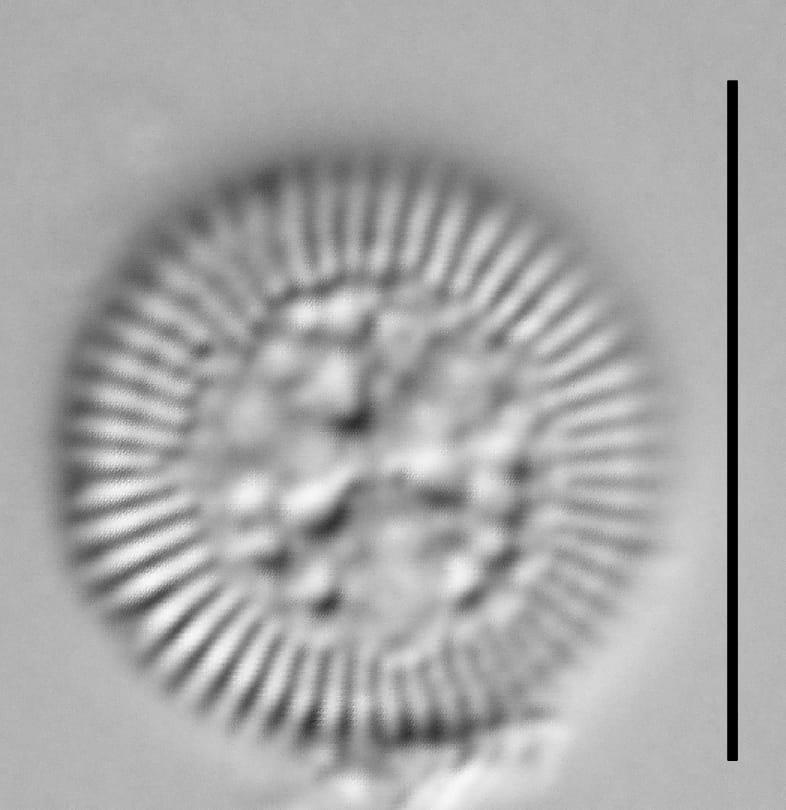
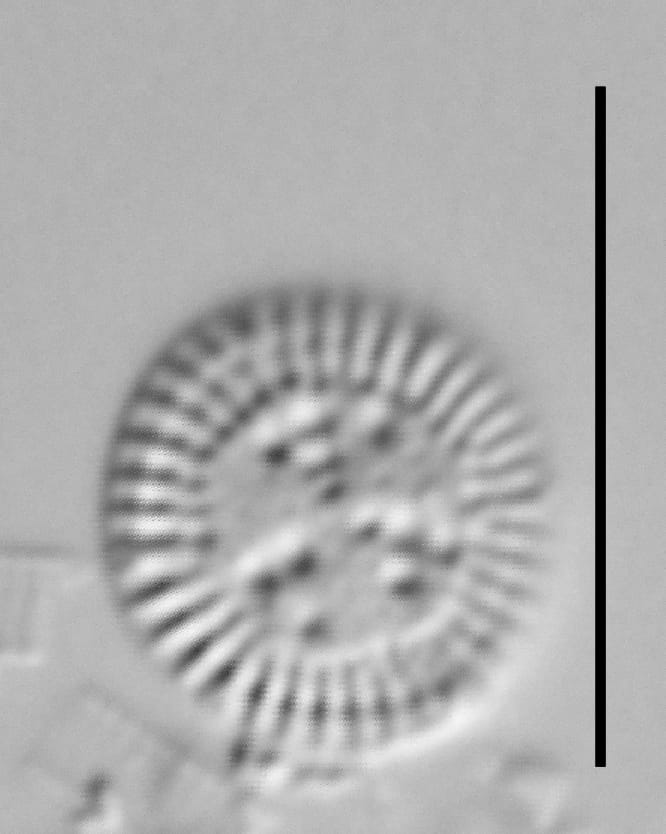
Valves are disc-shaped, with a low mantle and a flat valve face. The marginal area is narrow and composed of evenly spaced, fine alveolate striae extending approximately ⅓ to ½ the valve radius into the central area. The striae form a uniform radial pattern around the margin of the valve face, but the length of individual striae vary slightly on close inspection. A submarginal internal foramen is visible on each stria. The central area is indistinctly colliculate with ornamentation of variable size, number, and arrangement. Ornamentation as seen in the LM is a combination of two or more (2-3(5)) central fultoportulae (Houk et al. 2010), and pits or depressions that do not fully penetrate the valve. Ornamentation may form linear, well-defined rows at 120° angles, loosely clustered rows, wedge-shaped groups, or have a nearly random distribution across the valve face. Sierra Nevada populations commonly show ornamentation loosely clustered into 3 wedges. A single rimoportula is present near the perimeter of the central area, but it may be difficult to distinguish in LM. Marginal fultoportulae are present on every second to fifth costa, but are not easily seen in LM.
TAXONOMIC NOTES: Cyclotella is one of the more difficult diatom genera to conclusively identify specimens to the species-level as there is much confusion within the literature concerning species boundaries. Many species may be polymorphic, heterovalvar, or show biogeographical variability which is not accounted for in current species descriptions. The ecology of most species is poorly known. Additionally, it has been suggested that the structure of the central valve face is so variable as to be irrelevant to the identification process (Knie and Hübener 2007). Some workers (e.g. Tuebner 1995, Cremer et al. 2001) choose to designate populations that show great, but intergrading, variability as "complexes", such as the Cyclotella rossii-comensis-tripartita complex of Cremer (2001) or the Cyclotella ocellata-krammeri-rossii complex of Knie and Hübener (2007). Until significant work is undertaken to conclusively determine the nature of C. rossii, the taxonomic and ecological relevance of it morphotypes, similar information on its associated species, using the term 'Cyclotella-complex' or differentiating morphotypes may be the most practical approach.
Other taxa that appear closely allied to Cyclotella rossii include C. krammeri, which was designated as a valid name for the (potentially) invalid Cyclotella kuetzingiana sensu Grunow 1878 non Thwaites 1848, but excludes C. kuetzingiana var. planetophora Cleve-Euler and C. kuetzingiana var. radiosa Fricke in Schmidt. Others (e.g. Houk et al. 2010) continue to recognize C. kuetzingiana Thwaites and state that it is synonymous with C. krammeri. The central area of C. kuetzingiana/krammeri is slightly undulate and much less colliculate than C. rossii. Areolae and/or depressions show no pattern, or a weakly radiate pattern (sensu C. kuetzingiana var. radiosa).
Cyclotella comta var. oligactis Grunow in Van Heurck is probably conspecific with C. rossii, but Häkansson (1990) could not conclusively determine the nature of C. comta var. oligactis from available material and thus designated C. rossii a new species.
Cyclotella tripartita was described from Alaska and is considered synonymous with C. comensis sensu Manguin non Grunow (Håkansson 1990). The valve face ornamention of C. tripartita forms a trefoil pattern, very similar to the pattern observed in some members of our population of C. rossii. The valve face of C. tripartita, however, is radially undulate, not flat. Houk et al. (2010) suggest that C. tripartita may be conspecific with C. rossii based on similar, but unspecified, ecologies.
Cyclotella rossii was found in core sample from Fallen Leaf Lake, California, a deep, subalpine, meso-oligotrophic lake in the Tahoe Basin. Fallen Leaf Lake is composed of bicarbonate-type water with average pH 7.5-7.8, very low conductivity (8-12 ppm), and Secchi depth = 12 m (average). Cyclotella rossii can be found as a significant fraction of the plankton in the modern lake and is present as a subfossil to at least 3 m depth in the lake sediments, most likely representing several thousand years.
Reports of C. rossii in North America appear to be relatively limited; however, related species are common. Håkansson (1990) reports that C. rossii occurs in oligotrophic lakes with low conductivity. Cremer et al. (2001) report the species from East Greenland preferring water with low electrolyte content and pH around 7. However, based on core reconstructions, they suggest that C. rossii is not limited to oligotrophic lakes and may in fact be found in eutrophic conditions. Camburn and Charles (2000) report C. rossii from a shallow (24 m maximum depth) pond in the Adirondacks with conductivity = 28 μS/cm, pH 7.03, total alkalinity = 118 μeq/L, and Secchi depth = 6.0 m.
Many researchers note the need for significant work on the autecology of C. rossii, particularly as a method to conclusively determine valid species in the C. rossii complex; however, this challenge remains unmet.

Valve circular, with a diameter of 5-18 μm. Central part of the valve face flat, but sometimes slightly concentrically undulate. Central zone of the valve face with 2-5, seldom more, punctate, regular or irregular rows. Marginal zone narrow, regularly striate, but the length of the striae not always equal, without shadow lines. Marginal fultoportulae on every second or third interstria.


Transferred on this website from C. rossi to L. rossi, following Nakov et al. (2015). - S. Spaulding
Stratton, L. (2011). Lindavia rossii. In Diatoms of North America. Retrieved May 01, 2026, from https://diatoms.org/species/46613/cyclotella_rossii
The 15 response plots show an environmental variable (x axis) against the relative abundance (y axis) of Lindavia rossii from all the stream reaches where it was present. Note that the relative abundance scale is the same on each plot. Explanation of each environmental variable and units are as follows:
ELEVATION = stream reach elevation (meters)
STRAHLER = distribution plot of the Strahler Stream Order
SLOPE = stream reach gradient (degrees)
W1_HALL = an index that is a measure of streamside (riparian) human activity that ranges from 0 - 10, with a value of 0 indicating of minimal disturbance to a value of 10 indicating severe disturbance.
PHSTVL = pH measured in a sealed syringe sample (pH units)
log_COND = log concentration of specific conductivity (µS/cm)
log_PTL = log concentration of total phosphorus (µg/L)
log_NO3 = log concentration of nitrate (µeq/L)
log_DOC = log concentration of dissolved organic carbon (mg/L)
log_SIO2 = log concentration of silicon (mg/L)
log_NA = log concentration of sodium (µeq/L)
log_HCO3 = log concentration of the bicarbonate ion (µeq/L)
EMBED = percent of the stream substrate that is embedded by sand and fine sediment
log_TURBIDITY = log of turbidity, a measure of cloudiness of water, in nephelometric turbidity units (NTU).
DISTOT = an index of total human disturbance in the watershed that ranges from 1 - 100, with a value of 0 indicating of minimal disturbance to a value of 100 indicating severe disturbance.