Brachysira chiaruccii

-
Category
-
Length Range13.1-43.9 µm
-
Width Range4.5-6.8 µm
-
Striae in 10 µm29-32
-
Reported AsBrachysira procera (Bahls 2021, pg. 486, Fig 4-13)
Brachysira neoexilis morphotype 1 (Fallu et al. 2000, pg. 158, Pl. 12, Fig 11)
Brachysira microcephala morphotype 1 (Siver et al. 2005, pg. 251, Pl. 3, Fig 8)
-
ContributorLane Allen, Mark Edlund - Mar 2025
-
ReviewerBart Van de Vijver - Jun 2025
Identification






Description
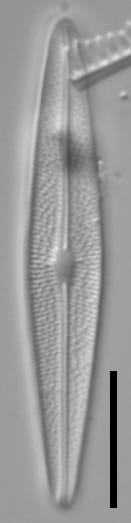
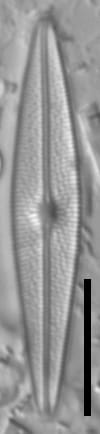
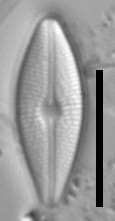
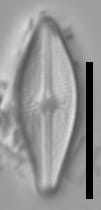
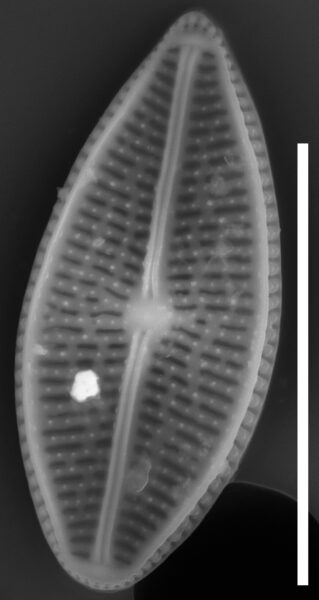
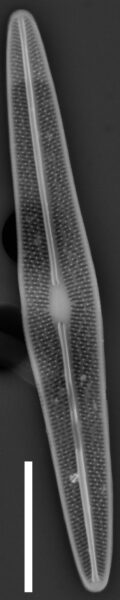
Valves are weakly heteropolar. The valve outline is lanceolate, with gently rounded apices. The footpole may be very weakly produced. Valve margins are weakly convex near the headpole, and even more convex in small specimens.
The axial area and central area are narrow. In large specimens, the central area is expanded along the apical axis, while in small specimens, the central area is indiscernible. The raphe is straight. Proximal raphe ends are straight but not are not expanded although they may be weakly deflected unilaterally in some specimens.
Striae are gently radiate near the central area, becoming more strongly radiate near the apices, becoming parallel at the apices. The angle of striation at the apices may be difficult to discern in LM. Voigt discontinuities are not apparent.
Externally, valves bear a modest marginal and axial crest. Small, rounded papillae are abundant on the valve face occurring on the virgae, forming a loosely decussate pattern. The distal raphe ends have an anchor shaped canal. Areolae are narrow and transapically expanded, with undulate margins. The adaxial areolae are typically the shortest, while those bordering the valve margin and central area are often the most elongate. Areolae are set in shallow depressions, as in Nupela.
Like many Brachysira species, pores on the mantle are aligned with a stria, except for the apices, where a series of notably narrower pores occur on the mantle. In this taxon, these pores extend slightly onto the valve face and wrap around the apices, typically extending slightly further around the footpole.
Internally, areolae bear hymens and small helictoglossae occur at the distal raphe ends.
Compère identified morphological diversity within the concept of B. microcephala, proposing two distinct morphotypes (Compère 1986, 1988). Morphotype 1 was defined by smaller apices (more closely aligned with B. microcephala s.s.), while mophotype 2 was defined by the distinctly capitate apices, aligning with B. neoexilis.
Autecology
This taxon has a circumboreal distribution. It is known to occur in oligotrophic, circumneutral, low conductivity lakes (Cantonati et al. 2021). Originally described from mountain lake sediments in Italy, this taxon has been documented with light micrographs from Québec (Fallu et al. 2000, as B. neoexilis morphotype 1), Cape Cod (Siver et al. 2005, as B. microcephala morphotype 1), a wetland in Montana (Bahls 2021, as B. procera), as well as Wisconsin (Apostle Islands).
In the Great Lakes National Parks (Edlund et al. 2009, 2011, 2012, 2013), this taxon is present in surface sediments of Stockton Lagoon (Apostle Islands) and rare on Isle Royale (Ahmik Lake).
Several research groups have used this taxon as an indicator organism and published regional environmental optima and tolerances including:
Fallu et al. (2000): Dissolved Organic Carbon - 4.52 +/- 1.70 mg/L for populations from Québec (as B. neoexilis morphotype 1).
Siver et al. (2005): pH - 5.65 +/- 0.65, Total Phosphorus (TP) - 10-1 +/- 4.7 µg/L for populations from Cape Cod (as B. microcephala morphotype 1).
-
Motility
-
Attachment
-
Habitat
-
Colony
-
Waterbody
-
Distribution
- Learn more about this
Original Description
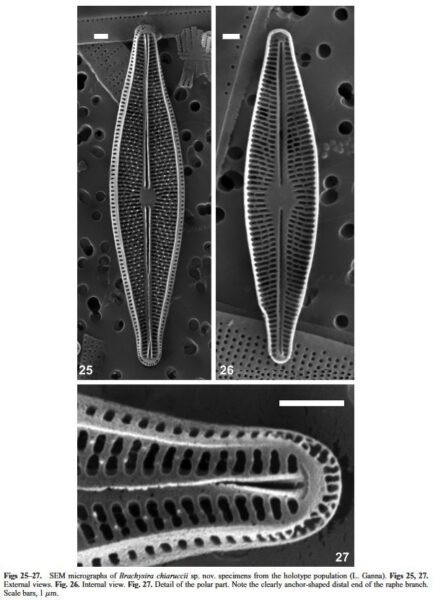
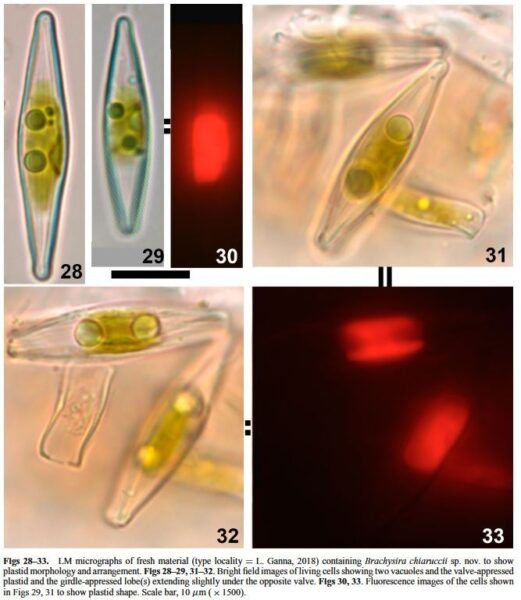
LM (Figs 1–24): Cells solitary. Frustules rectangular to narrowly rectangular in girdle view, with two to several girdle bands. Valves narrowly rhombic to lanceolate, often slightly heteropolar, tapering uniformly to weakly or barely protracted, acutely rounded apices, that sometimes become slightly subcapitate in the smallest specimens. Valve dimensions: length 14.0–40.0 μm, width 4.0–6.5 μm, length:width 3.5–7.5 (Table 2). Axial area narrowly linear, only widening very close to the central area. Central area narrowly elliptic to elliptic in smaller specimens. Raphe straight, filiform with simple, straight, proximal raphe endings. Striae slightly radiate to almost parallel at centre, 32–35 in 10 μm. Voigt discontinuity, and apical alignment of areolae not visible. SEM External view (Figs 25–27): Valve face flat, sur- rounded by a mostly corroded, faint ridge (Fig. 25). Distal raphe endings distinctly anchor shaped. Raphe branches bordered by strong ribs (Figs 56, 57). Striae uniseriate, slightly radiate to almost parallel at centre, parallel to weakly convergent at apices. Striae composed of 1–3 transapically elongated areolae, similar or dissimilar in length; 2–5 shortened striae bordering the central area. Small pointed papillae present on virgae, 1–4 per interstria (single papilla at apices). Single elongate areola, lacking papillae on mantle, closer together around apices. Areola foramina with small indentations. SEM Internal view (Fig. 26): Proximal raphe ends only very-slightly bent, raphe branches terminating in faint helictoglossae. As it is typical for the genus, a single, valve-appressed plastid with girdle-appressed lobe(s) extending slightly under the opposite valve. Two prominent vacuoles of variable size, typically seen in living cells (Figs 28–33).
Holotype. cLIM005 DIAT 3513 (Diatom collection of the MUSE – Museo delle Scienze, Trento, Italy, Herbarium: TR) (Registro CRAD 2016162; Lago di Ganna, periphyton on submerged reed stems). Collected by A. Beghi and F. Pandolfi on the 8th of July 2016
Isotypes. BM 81901, Diatom Collection of the Natural History Museum (BM), London, UK; B 40 0045145, Diatom collection of the Botanischer Garten und Botanisches Museum Berlin-Dahlem, Zentraleinrichtung der Freien Universität Berlin, Germany (B).
Registration. http://phycobank.org/102896
Type locality. Lake Ganna (Lago di Ganna, Italy; Table 1).
Etymology. The species is dedicated to Prof. Alessandro Chiarucci, University of Bologna, for his outstanding contributions to vegetation ecology, and his efforts to bolster the development and innovation of plant sciences in Italy. Comparison with similar species. Some populations of Brachysira neoexilis Lange-Bertalot (isopolar) may be similar, but this species has consistently rostrate to (sub)capitate endings. Similarly B. neglectissima Lange-Bertalot (isopolar) has more or less protracted ends, a roundish-rhombic central area, and is smaller (in particular shorter) (Table 2). Brachysira conamarae Kennedy et Allott (isopolar) has a higher stria density (34–37 in 10μm) and an indistinctly lanceolate central area, which is only slightly wider than the axial area. Brachysira liliana Lange-Bertalot (isopolar) has a higher stria density (36–40 in 10 μm), and a narrowly elliptical central area (Table 2). Brachysira procera Lange-Bertalot et Gerd Moser (isopolar) has a lower stria density (27–30 in 10 μm), and a less developed central area. The heteropolar Brachysira ocalanensis H.A. Shayler et P.A. Siver has a higher stria density (33–43 in 10 μm), and more narrowly lanceolate outline with more acutely rounded apices. Brachysira ontonageniana Kociolek et Lowe, also heteropolar, has less acute poles.
Distribution. Not yet known because the species has previously not been distinguished from other Brachysira taxa. So far it is only known from the type locality (Lake Ganna, northern Italy), and another mountain lake, Lake Ninu in Corsica (Figs 34–58). The substrata sampled in the two lakes were submerged Phragmites stems (40–50 cm depth) and phytoplankton collected above the lake bottom, respectively. Maximum relative abundance was 9.2% in Lake Ganna and 8.5% in Lake Ninu.
Ecology & co-occurring diatom species. The locus classicus (Lake Ganna) is an oligotrophic, shallow lake, rich in macrophytes, in particular Nymphaea alba L. and Schoenoplectus lacustris (L.) Palla, with relatively low conductivity (about 100 μS cm−1 ), the euphotic zone extending throughout the entire vertical profile (Table 1). This moderate elevation lake differs in several features (Table 1) from the vegetation-poor, mountain, oligo-mesotrophic, deep, dimictic Lake Ninu (Loye-Pilot et al. 1997). The most frequent and abundant diatom species occurring together with the new species at the type locality were Tabellaria flocculosa (Roth) Kützing (37.7%), Achnanthidium minutissimum (Kützing) Czarnecki (19%), Ulnaria delicatissima (W.Smith) Aboal et Silva (5.2%), Encyonopsis cesatii (Rabenhorst) Krammer (3.9%), Cymbella cymbiformis Agardh (2.9%), Gomphonema lateripunctatum Reichardt et Lange-Bertalot (2.2%). Brachysira neoexilis and B. styriaca were also present in the sample. In Lake Ninu 49 diatom taxa were identified, and the community was dominated by Brachysira, Eunotia, Fragilaria, and Tabellaria. The most frequent and abundant species occurring together with the new species were Fragilaria gracilis Østrup (23%), Tabellaria flocculosa (14.6%), B. neoexilis (14.4%), Eunotia spp. C.G. Ehrenberg (7.4%); the subdominant species were Achnanthidium minutissimum (4.7%), Pseudostaurosira brevistriata (Grunow in Van Heurck) Williams et Round (3.2%), Achnanthidium neomicrocephalum Lange-Bertalot et F. Staab (2.7%), and B. neglectissima (1.9%).
-
AuthorCantonati, Lange-Bertalot, Arnaud, Galbiati and Soróczki-Pintér 2021
-
Length Range14.0-40.0 µm
-
Width3.5-7.5 µm
-
Striae in 10µm32-35
Original Images


Citations & Links
Citations
-
Publication Link: 10.1080/0269249X.2021.1982778
-
Publication Link: 10.1080/23818107.2021.1909499
-
Publication Link: 10.1127/nova_hedwigia/2023/0839
Links
-
Index Nominum Algarum
Cite This Page
Allen, L., Edlund, M. (2025). Brachysira chiaruccii. In Diatoms of North America. Retrieved July 22, 2026, from https://diatoms.org/species/341255/brachysira-chiaruccii
Responses
The 15 response plots show an environmental variable (x axis) against the relative abundance (y axis) of Brachysira chiaruccii from all the stream reaches where it was present. Note that the relative abundance scale is the same on each plot. Explanation of each environmental variable and units are as follows:
ELEVATION = stream reach elevation (meters)
STRAHLER = distribution plot of the Strahler Stream Order
SLOPE = stream reach gradient (degrees)
W1_HALL = an index that is a measure of streamside (riparian) human activity that ranges from 0 - 10, with a value of 0 indicating of minimal disturbance to a value of 10 indicating severe disturbance.
PHSTVL = pH measured in a sealed syringe sample (pH units)
log_COND = log concentration of specific conductivity (µS/cm)
log_PTL = log concentration of total phosphorus (µg/L)
log_NO3 = log concentration of nitrate (µeq/L)
log_DOC = log concentration of dissolved organic carbon (mg/L)
log_SIO2 = log concentration of silicon (mg/L)
log_NA = log concentration of sodium (µeq/L)
log_HCO3 = log concentration of the bicarbonate ion (µeq/L)
EMBED = percent of the stream substrate that is embedded by sand and fine sediment
log_TURBIDITY = log of turbidity, a measure of cloudiness of water, in nephelometric turbidity units (NTU).
DISTOT = an index of total human disturbance in the watershed that ranges from 1 - 100, with a value of 0 indicating of minimal disturbance to a value of 100 indicating severe disturbance.
Brachysira chiaruccii
- Valves weakly heteropolar
- Apices acutely rounded to weakly produced
- Striae discernable in LM
This taxon is weakly heteropolar; the headpole is acutely rounded and the footpole is weakly produced. Stria and areolae are readily visible in LM. Striae are radiate. Valve margins are weakly convex.