
Credit: WPLG Miami
Aerial view of Chaetoceros spp. Bloom, dominated by C. lauderi. Biscayne Bay, Miami, Florida, 21 August 2020






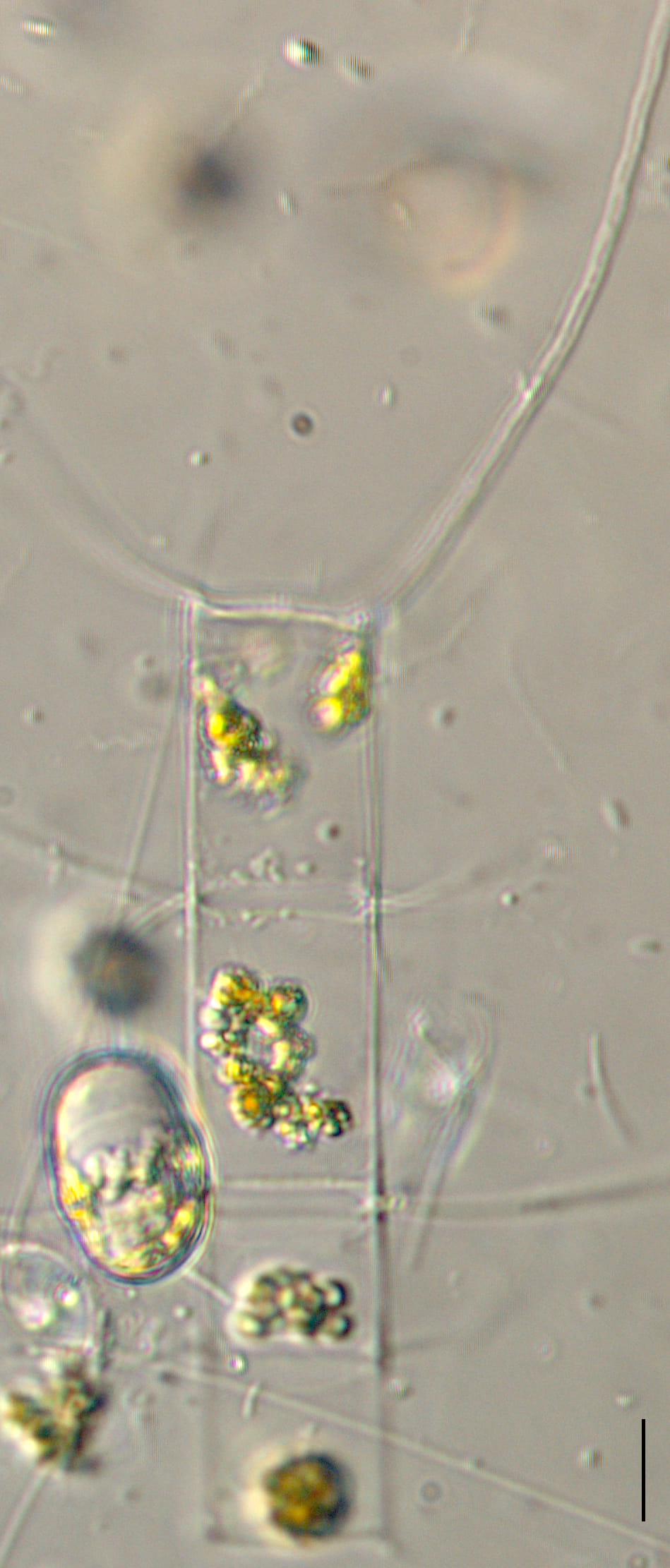
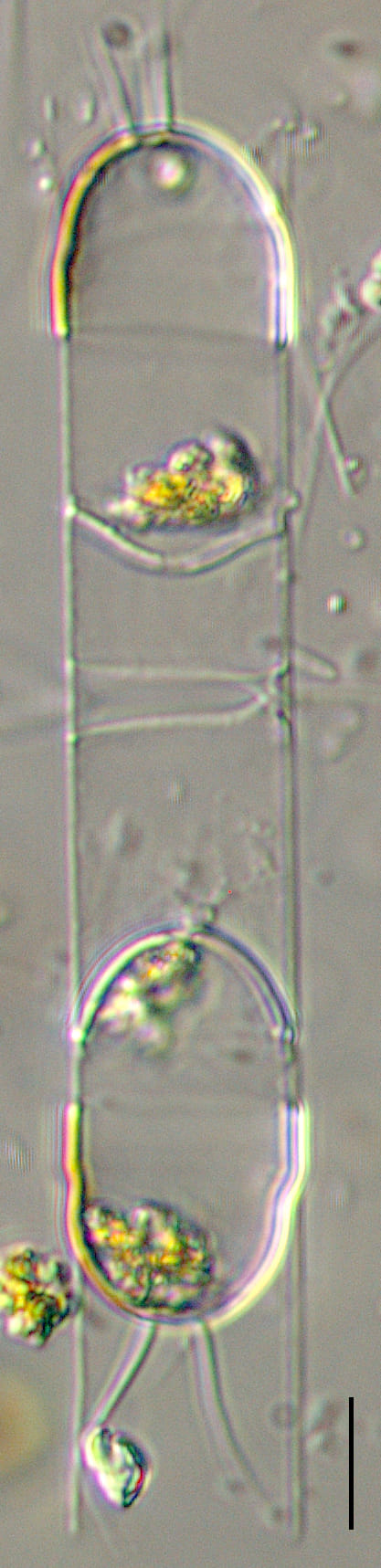
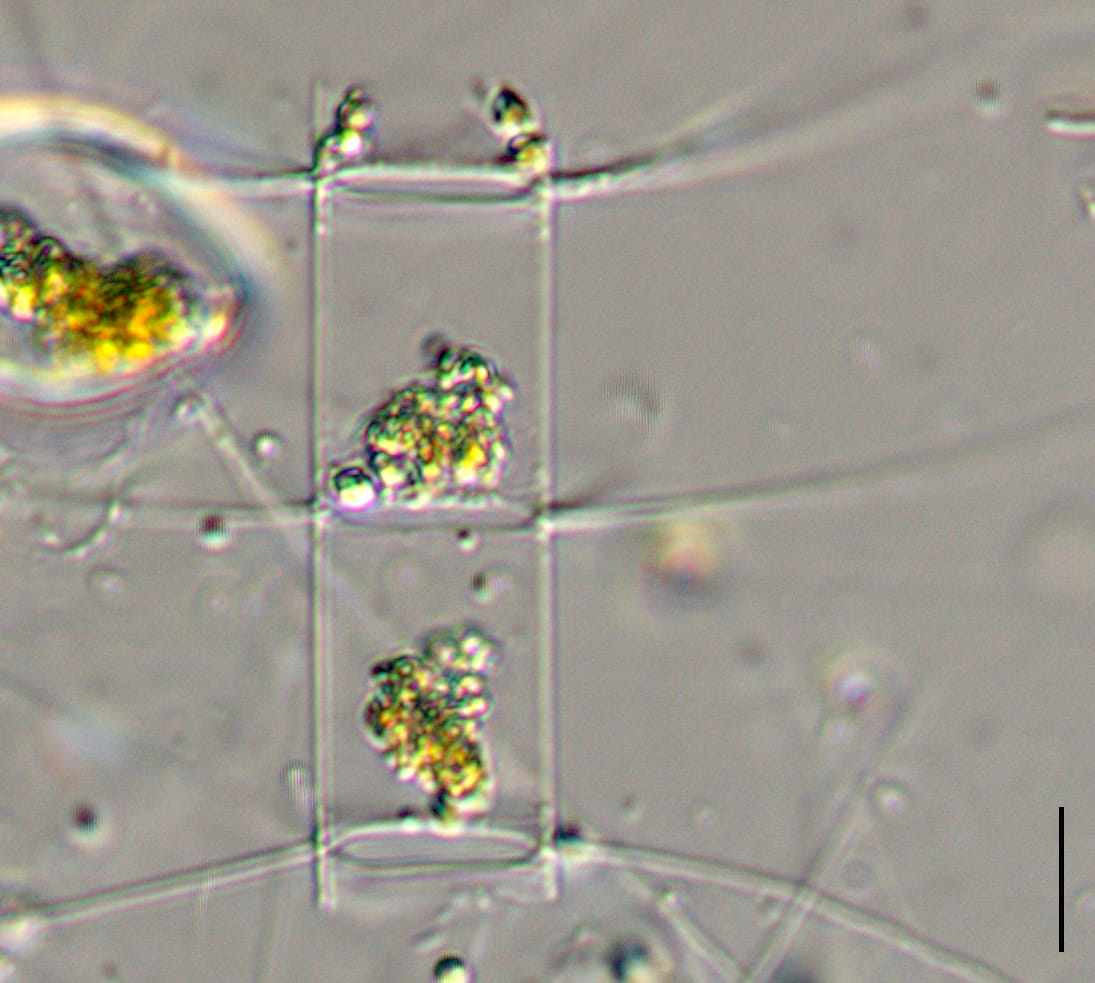
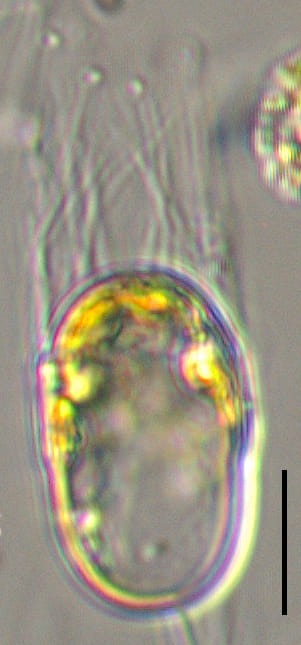
Cells are joined together in colonies of medium length (4-10 cells). Frustules are lightly silicified and separate upon drying. In girdle view, cells appear to be rectangular, with “sharp corners” and narrow linear apertures. The pervalvar axis is longer than the apical axis, in most specimens. The suture between the valve mantle and the cingulum is indistinct.
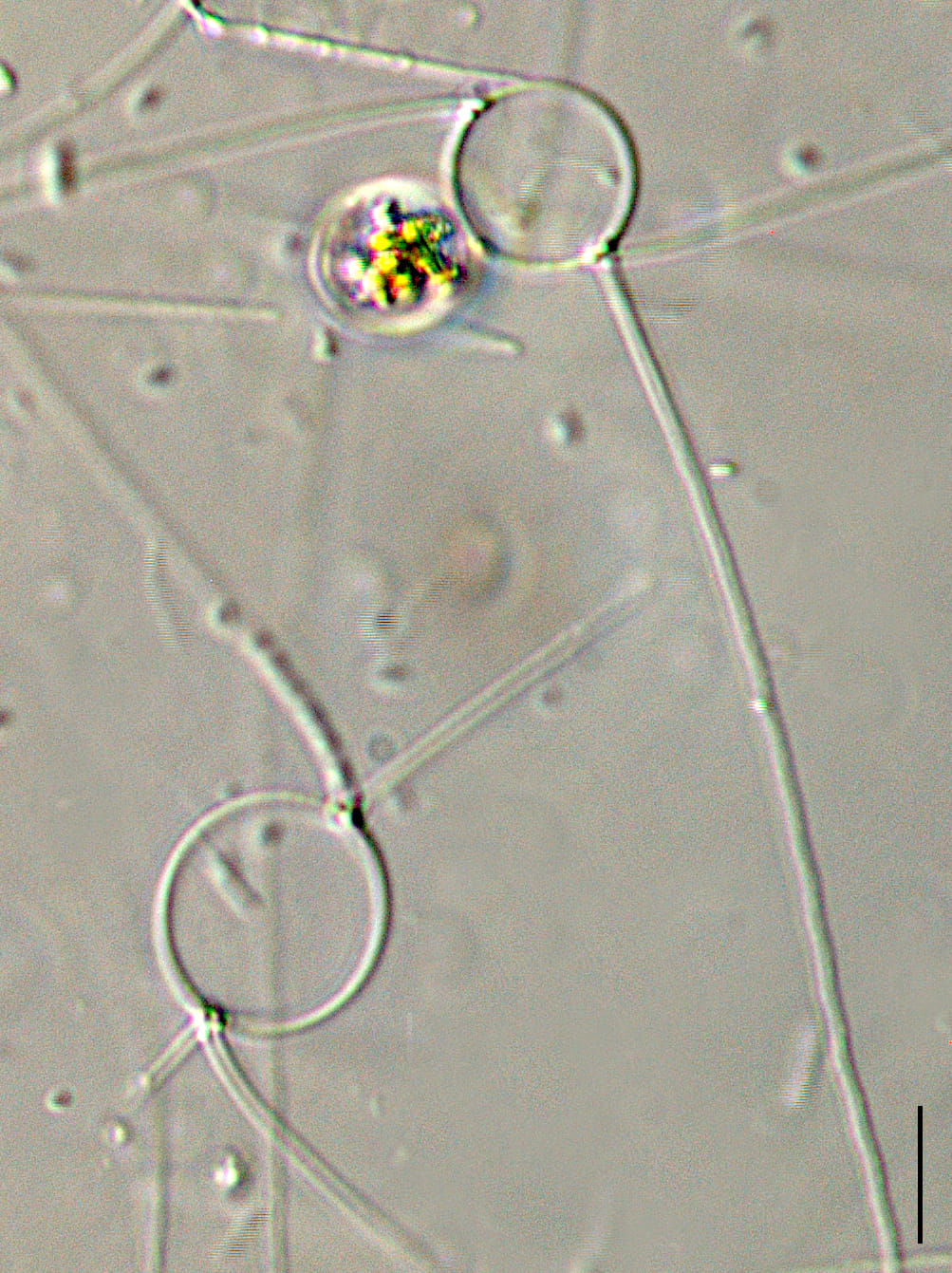
Setae are long (> 150 microns) and oriented nearly perpendicular to the chain axis. Terminal setae are similar to intercalary setae in thickness and length. The terminal setae emerge parallel to the apical axis and then bend towards the chain end but remain divergent over their entire length. Intercalary setae are fused at the plane of the valve margin.
The valve face is flat. Valves are nearly circular in outline. Chloroplasts are numerous, small, and confined within the main body of the cell (no chloroplasts are present within the setae). Resting spores are formed within an elongated mother-cell. The primary valve of each resting spore is elevated from the valvar plane, hemispherical and sometimes “doorknob” shaped, and has long, sparse spine(s). The secondary valve of each resting spore is less elevated and spines may be present or absent on the valve face. Secondary valves also have a curtain of variously-developed capillate spines extending from the advalvar margin.
In SEM, valves were observed in valve view to have partially collapsed, shallow mantles. Setae are circular in cross section with spirally arranged rows of closely spaced poroids and conical spines. The resting spore morphology is variable, with primary valves varying from hemispherical to "doorknob" in shape. Spines are sparse relative to the original description (Ralfs ex Lauder 1864), but fall within the range of variation observed in the species (Paul Hargraves and Jan Rines, personal email communications - Feb. 2021). Primary valves of resting spores with radial arrangement of closely spaced granulae and sparse relatively long spines.
The genus Chaetoceros is species-rich, and for most species there is often not a single diagnostic character to distinguish taxa. Rather, a unique combination of character states distinguish taxa. Determining these character states will help the practitioner to identify Chaetoceros specimens to one of 15 monophyletic Sections (De Luca et al. 2019) or to undescribed taxa.
For C. lauderi, it is important to verify the presence of multiple small plastids confined to main body of the cell, rather than multiple plastids positioned within the main body and setae (Section Chaetoceros) or a few larger plastids within the main cell body (most other Sections). Therefore, live material or preserved cells are necessary. The presence of these character states indicates a much smaller subset of Chaetoceros species (belonging to 3 Sections). Most Chaetoceros species have one or two larger plastids, or smaller plastids found throughout the cell, including the hollow setae.
Observation of narrow apertures between cells in girdle view and nearly circular valve outlines in valve view is indicative of members of the Section Cylindrica, consisting of two species (C. lauderi and C. teres).
Distinguishing these two taxa requires examination of the primary valves of
the resting spores. The primary valve face of C. lauderi is spinose,
while that of C. teres lacks spines. Ultrastructure of the setae is
also taxonomically informative (see above). The setae of C. teres are
generally shorter (about one half the length) and finer. Finally, definitive
determination between the two species requires the examination of resting
spores.
Chaetoceros lauderi is a planktonic diatom and has been documented throughout the world in marine and estuarine tropical to temperate nearshore waters in the Atlantic: Narragansett Bay (Rines and Hargraves 1988), Puerto Rico (Navarro 1982), Adriatic Sea (Bosak and Sarno 2017), Baltic Sea (Hällfors 2004), Black Sea (Barinova et al. 2011), Brazil coast (Eskinazi-Leça et al. 2010); and Pacific Oceans: California coast (Cupp 1943), Mexico coast (Torres-Ariño et al. 2019), Sea of Japan (Shevchenko et al. 2019), and Australia coast (McCarthy 2013).
Based on global distribution and seasonal occurrence, C. lauderi is a warm water diatom, while the similar C. teres is more of a cold water species.

C. Lauderi, n. sp., Ralfs. Filaments with quadrate frustules, narrow-oblong fenestrae, slightly and gradually constricted at the middle, and long awns; sporangia with very unequal, spinous valves, the larger one capitate (fig. 4). Hong Kong.
About the middle of April, and shortly previous to the disappearance of the filaments, sporangia are seen, in which the large valve is capitate, hirsute, and connected to the hoop by a broad, smooth neck. The smaller conical valve is furnished with spines, or a delicate plicated membrane resembling spines. I think the latter, because there is no trace of this lower part with membrane attached after boiling in acid. There is also a fine plicated membrane attached to the margins of the hoop. I think it is a large variety without the constricted neck. Breadth of a, 1/900”.


Frankovich, T. (2020). Chaetoceros lauderi. In Diatoms of North America. Retrieved July 22, 2026, from https://diatoms.org/species/96737/chaetoceros-lauderi
The 15 response plots show an environmental variable (x axis) against the relative abundance (y axis) of Chaetoceros lauderi from all the stream reaches where it was present. Note that the relative abundance scale is the same on each plot. Explanation of each environmental variable and units are as follows:
ELEVATION = stream reach elevation (meters)
STRAHLER = distribution plot of the Strahler Stream Order
SLOPE = stream reach gradient (degrees)
W1_HALL = an index that is a measure of streamside (riparian) human activity that ranges from 0 - 10, with a value of 0 indicating of minimal disturbance to a value of 10 indicating severe disturbance.
PHSTVL = pH measured in a sealed syringe sample (pH units)
log_COND = log concentration of specific conductivity (µS/cm)
log_PTL = log concentration of total phosphorus (µg/L)
log_NO3 = log concentration of nitrate (µeq/L)
log_DOC = log concentration of dissolved organic carbon (mg/L)
log_SIO2 = log concentration of silicon (mg/L)
log_NA = log concentration of sodium (µeq/L)
log_HCO3 = log concentration of the bicarbonate ion (µeq/L)
EMBED = percent of the stream substrate that is embedded by sand and fine sediment
log_TURBIDITY = log of turbidity, a measure of cloudiness of water, in nephelometric turbidity units (NTU).
DISTOT = an index of total human disturbance in the watershed that ranges from 1 - 100, with a value of 0 indicating of minimal disturbance to a value of 100 indicating severe disturbance.