Eolimna metafarta

-
Category
-
Length Range11.4-23.3 µm
-
Width Range6.7-8.9 µm
-
Striae in 10 µm11-13
-
Reported AsNavicula farta (Foged 1981, pg. 262, Pl. 37, Fig 4)
-
ContributorLane Allen, Mark Edlund - May 2025
-
ReviewerSarah Spaulding - May 2025
Identification




Description
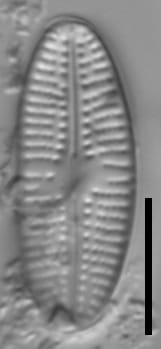
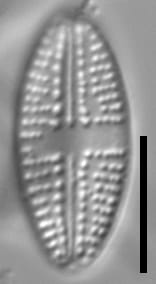
This taxon has robust valves, with a flat valve face and broadly elliptic margins. Apices are broadly rounded. Striae are very coarse and parallel to slightly radiate at the center of the valve, becoming slightly more radiate at the apices. The adaxial areolae appear larger and more distinct, forming a row on both sides of the raphe. There are typically small interruptions in the striae between the adaxial areolae and the rest of the areolae in most striae. As a result of the absence of adaxial areolae, the central area is poorly defined, with striae approaching the central area to varying degrees. The striae are often irregularly spaced bordering the central area. In some specimens (uncommon), the central area may extend to the valve margin on one side. The areola density (excluding the adaxial areolae) is approximately 15-20 in 10 µm and should be resolvable in LM.
The raphe is typically straight, although may be slightly arcuate in small specimens. The raphe is simple, tapering slightly at the apices. The proximal raphe ends may be slightly expanded, and may be weakly deflected unilaterally. The distal raphe ends are strongly deflected unilaterally in the opposite direction of the proximal raphe ends. The axial area tapers near the apices.
This taxon is one of the larger species in the genus, for example, Kulikovskiy et al. (2015) reported specimens up to 33 µm long from Lake Baikal.
This taxon was originally illustrated by Hustedt in Schmidt's Atlas (1934); however, at that time, a written description was not provided, invalidating the name. Hustedt referenced this taxon again in 1952, but failed to validate the name. Hustedt published a formal description in 1966, but did not include a description in Latin, which at that time, rendered that name invalid still. In 2015, Kulikovskiy et al. encountered a taxon that appeared to be a match for Hustedt's taxon, but were uncertain if their taxon was in fact the same species as Hustedt's taxon. Recognizing that this taxon does not belong in the genus Navicula, Kulikovskiy et al. (2015) described Eolimna metafarta, hesitantly proposing a synonomy with Navicula farta Hustedt. More work is required to determine if the population investigated by Hustedt and the population investigated by Kulikovskiy et al. (2015) belong to the same species.
From August 2019 to May 2025, a different and possibly undescribed taxon currently recognized as Eolimna sp.1 SESQA was identified as Eolimna metafarta. Populations identified as E. metafarta based upon that morphological species concept should be re-evaluated. Check the update for Eolimna sp.1 SESQA for more information.
Autecology
This taxon is rare with a disjunct distribution within circumboreal regions (Hustedt 1934, Foged 1981, Kulikovskiy et al. 2015). In North America, this taxon has been documented in lakes based on light micrographs from Alaska (Foged 1981), Montana, Idaho (Bahls 2023), and Michigan (this page).
In the Great Lakes National Parks (Edlund et al. 2009, 2011, 2012, 2013), Eolimna metafarta (reported as Navicula farta) is rare (~1% relative abundance) in sediment core material from Voyageurs (Mukooda Lake) and Pictured Rocks (Grand Sable and Beaver lakes) National Parks.
This taxon is rarely reported and appeared as a trace component of the community in two sites in North America, Grand Sable Lake, Michigan (this page) and Upper Priest Lake, Idaho (Bahls 2023). Because of it's rare occurrence, there are no estimates of its autecology. However, this taxon co-occurred with the other rare and distinctive taxa, Gomphosphenia grovei, Sellaphora schadei, Diploneis marginestriata, and Fallacia subhamulata in the two documented localities.
-
Size Range, µm3
-
Motility
-
Attachment
-
Habitat
-
Occurrence
-
Waterbody
-
Distribution
- Learn more about this
Original Description
Eolimna metafarta Kulikovskiy & Lange-Bertalot sp. nov. (Figs 1-54, 59-68)
Synonym: ?Navicula farta Hustedt in Schmidt et al. (1934: figs 397: 7-9), nom. inval, Hustedt(1966: 644, fig. 1646), nom. inval. (but see Simonsen 1987: 166-167)
GENERAL DESCRIPTION: LM observations (Figs 1-54), valves elliptical to linear-elliptical; ends broadly rounded. Length 8-33 um, breadth 4.5-10.5 um. Raphe filiform, straight or slightly curved with inconspicuous, barely deflected central ends and bent terminal fissures. Axial area variable in width, narrow and almost linear in smaller cell cycle stages but wider and more or less lanceolate in later stages. Central area variable in size, shape and symmetry; some specimens unilaterally extend almost to valve margin. Lateral area developed in larger and medium-sized specimens. Transapical striae radiate throughout, moderately so in proximal parts, becoming more radial distally, 11-13 in 10 um. Adaxial areolae of each stria form an apically aligned row on either side appearing more accentuated than others from which they are separated by the more or less distinct lateral area. The remaining areolae are somewhat irregularly more densely spaced, ca. 15 In 10 um.
SEM observations, external valve view (Figs 59-64), the raphe lies in an elevated sternum flanked by depressions that comprise the adaxial series of areola foramina. The raphe slit is not expanded, or only weakly expanded toward the proximal ends; terminal fissures both bent unilaterally, but to the opposite side of the slightly deflected proximal parts of the raphe. The relief-like appearance of the entire valve face concerns likewise the lateral areae being elevated between adaxial depressions and marginal parts of the valve. Sternum and lateral areae are connected on the same level by the "central area". Marginal parts characterized by striae that are composed of approximately circular uniseriate foramına lying in depressions between elevated virgae. Isolated areolae are generally absent on the mantle, but in some cases few ones are distributed at the ends.
Internal view (Figs 65 – 68), raphe slit straight, lying in the broad, slightly elevated internal sternum; central ends extremely weakly expanded toward central pores, slightly deflected. Apical ends with small helictoglossae. Areola apertures impressed into the surface, occlusions (here slightly dissolved) in a middle position between external and internal apertures. Valve mantle shallow, not perforated.
-
AuthorKulikovskiy and Lange-Bert. 2015
-
Length Range8-33 µm
-
Width4.5-10.5 µm
-
Striae in 10µm11-13
Original Images




Citations & Links
Citations
Cite This Page
Allen, L., Edlund, M. (2025). Eolimna metafarta. In Diatoms of North America. Retrieved July 24, 2026, from https://diatoms.org/species/342596/eolimna-metafarta
Responses
The 15 response plots show an environmental variable (x axis) against the relative abundance (y axis) of Eolimna metafarta from all the stream reaches where it was present. Note that the relative abundance scale is the same on each plot. Explanation of each environmental variable and units are as follows:
ELEVATION = stream reach elevation (meters)
STRAHLER = distribution plot of the Strahler Stream Order
SLOPE = stream reach gradient (degrees)
W1_HALL = an index that is a measure of streamside (riparian) human activity that ranges from 0 - 10, with a value of 0 indicating of minimal disturbance to a value of 10 indicating severe disturbance.
PHSTVL = pH measured in a sealed syringe sample (pH units)
log_COND = log concentration of specific conductivity (µS/cm)
log_PTL = log concentration of total phosphorus (µg/L)
log_NO3 = log concentration of nitrate (µeq/L)
log_DOC = log concentration of dissolved organic carbon (mg/L)
log_SIO2 = log concentration of silicon (mg/L)
log_NA = log concentration of sodium (µeq/L)
log_HCO3 = log concentration of the bicarbonate ion (µeq/L)
EMBED = percent of the stream substrate that is embedded by sand and fine sediment
log_TURBIDITY = log of turbidity, a measure of cloudiness of water, in nephelometric turbidity units (NTU).
DISTOT = an index of total human disturbance in the watershed that ranges from 1 - 100, with a value of 0 indicating of minimal disturbance to a value of 100 indicating severe disturbance.
Eolimna metafarta
- Adaxial areolae prominent
- Striae coarse
- Central area variable
This taxon distinguished by its coarse striae and prominent adaxial areolae. Valves are broadly elliptic, with broadly rounded apices. While the adaxial areolae are very regular, the rest of the striae are somewhat irregular.